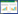 图 1 群落水生植物的相对生长速率
Fig.1 Relative growth rate of community aquatic plants
图 1 群落水生植物的相对生长速率
Fig.1 Relative growth rate of community aquatic plants
HONG Shuchen,WANG Chao'er,FENG Zhaorui,et al.Differential study on nitrogen and phosphorus accumulation in different aquatic plant communities[J].Journal of Zhejiang University of Science and Technology,2021,33(05):432-440.

水体富营养化是近年来中国地表水面临的普遍性污染现象,大量的氮、磷等营养元素伴随生活污水排入各种水体,导致藻类及其他浮游生物过量繁殖,水体中溶解氧含量下降,水质趋于恶化[1-3]。利用水生植物吸收水中氮、磷物质,具有较好的生态效应[4-7],因此成为地表水体修复的重要方法。当前利用水生植物修复水体的研究多集中于某一类型植物[8-10],而实际水环境中的植物类型众多,即植物在水体中是以群落层级存在,就文献而言,目前针对于群落植物修复水体的研究较为少见,相关认识亦不成熟,且缺少相应的实践活动。从种间关系分析,同一水环境中不同类型植物可能存在竞争也可能存在互补[11],如沉水植物多生长在水体中、下部,其对水体中游离态营养元素具有较强的富集效应; 然而挺水植物根系发达且可深入水底泥沙层,故可富集底泥中的营养元素[12-13]。显然,这两类植物类型的组合存在协同效应,但同时又由于各自生长习性的差异,也存在竞争关系[14-16]。
为了探究水生植物群落在不同水质条件下的生长状况及其对氮、磷的富集状况,宜选用美人蕉、鸢尾等氮、磷富集能力强的植物[17],同时还应考虑植物高低搭配以及水面及水下污染去除等要求,故研究中选择美人蕉(Canna indica)、鸢尾(Iris tectorum)作为高大型挺水植物,以梭鱼草(Pontederia cordata)、黑麦草(Lolium perenne)作为中等型挺水植物,以狐尾藻(Myriophyllum verticillatum)、铜钱草(Hydrocotyle chinensis)作为低矮浮水植物进行组合[18]。鉴于氮、磷为主要富集目标[19],故以美人蕉、鸢尾作为组群基础植物,分别与1种中等型及1种低矮型植物进行群落构建。通过水培试验,对比不同植物群落的生长状况及氮磷富集状况的差异,并就相关机理进行分析讨论,以期为植物群落用于富营养化水体的生态化治理提供合理的理论解释和相应的数据支持。
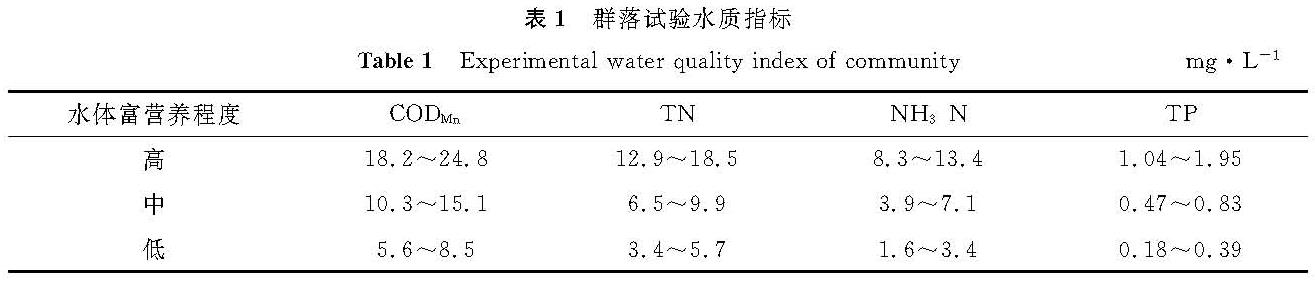
1 材料与方法1.1 试验概况试验在浙江省废弃生物质循环利用重点实验室基地大棚内进行,试验周期为2018年3月至2020年10月。所选取的6种植物包括不同挺水植物与沉水植物,取幼株种植在一起构成植物群落,植物按初始生物量大致相等原则构建群落,其中包括由鸢尾、黑麦草、美人蕉和狐尾藻组成的群落1,由鸢尾、黑麦草、美人蕉和铜钱草组成的群落2,由鸢尾、美人蕉、梭鱼草和铜钱草组成的群落3,由鸢尾、美人蕉、梭鱼草和狐尾藻组成的群落4。植物幼株取自杭州市区各河道,其中铜钱草取自西蜜湖上游河道,美人蕉、鸢尾、黑麦草、梭鱼草取自杭州西溪湿地,狐尾藻取自余杭塘河。同种植物选取幼株株高、外形基本上相近者作为待测对象,先用自来水将植株上的泥土清洗干净,再用蒸馏水清洗后待用。植物采用盆栽方式种植于瓦缸内,挺水类植物置于泡沫带孔板中固定,根部浸没于水中,藻类植物于水体中悬浮; 为进行合理对比,各植物初始生物量统一设定为(30±1.25)g干质量。以沟渠水浓缩液作为贮备液,自来水稀释后作为试验用水,试验水样体积维持在(65±0.5)L,试验期间损失的蒸发量根据液面刻度线补水并同时以磷酸氢二钾和氯化铵溶液进行氮、磷调节,以维持试验水量和氮、磷含量基本上稳定。群落试验水质配制同单一植物试验,水样体积为(65±0.5)L,试验期间损失的蒸发量根据液面刻度线补水并同时进行氮、磷调节,以维持试验水量和氮、磷含量基本上稳定。试验水质分别设置为高、中、低3种不同营养水平以测试各群落植物生长适应性及氮、磷富集能力差异。不同试验水质的化学需氧量(以高锰酸钾指数(CODMn)计)及总氮(TN)、氨氮(NH3-N)、总磷(TP)的含量见表1。
表1 群落试验水质指标
Table 1 Experimental water quality index of communitymg·L-1
1.2 分析方法
净增生物量:用吸水纸擦去植株表面水,用电子天平称其鲜质量,之后在105 ℃烘箱杀青1 h,再于80 ℃烘箱中烘至恒重后称其干质量,净增生物量以试验开始及试验结束(包括试验过程中掉落叶片等组织)时的干质量差计。
氮、磷含量:将烘干后的植物样品研磨成粉,过100目筛后装瓶保存; 定量称取样品,采用浓硝酸-高氯酸消解法进行预处理,消解后取上清液,定容后待测。TN、TP分别采用HJ 636—2012《水质 总氮的测定 碱性过硫酸钾消解紫外分光光度法》[20]和HJ 671—2013《水质 总磷的测定 流动注射-钼酸铵分光光度法》[21]测定。
相对生长速率(relative growth rate,RGR)

式(1)中:St、S0分别为试验结束时和试验开始时的生物量,g; t为水生植物生长时间,取50 d。
积累率
X=(Mt-M0)/(M0)。 (2)
式(2)中:Mt、M0分别为试验结束时和试验开始时各水生植物中氮(磷)的含量,g。
2 结果与分析2.1 群落中各水生植物生长状况各群落水生植物生长状况见图1,图中不同小写字母表示不同营养水平下的同种植物在0.05水平下具备显著性差异; 图例L、M、H分别表示低、中、高3种营养水平(文中以下部分同)。试验结果显示出同一水生植物在不同群落中的生长状况存在差异,同一群落中的水生植物在不同营养条件下的生长状况也不尽相同,说明群落中的不同水生植物间存在相互影响。
由图1(a)可知,群落1中4种植物随营养水平的提高,均呈现出生长速率逐渐增大的趋势。其中,鸢尾在3种营养水平下的增长速率均存在显著性差异,其余3种植物在低、高营养水平下的增长速率均有显著性差异,但黑麦草、狐尾藻两种植物在中营养水平下的增长速率均与低、高营养水平下的增长速率无显著性差异,美人蕉在低、中营养水平下存在显著性差异,在中、高营养水平下无显著性差异。从整体上看,美人蕉在3种营养水平下的增长速率均最大,而狐尾藻的增长速率则最小。由图1(b)可以看出,美人蕉在3种营养水平下的增长速率均高于群落2中其他植物,且呈现出营养水平间的显著性差异; 鸢尾与黑麦草增长速率与黑麦草相近,但在低营养水平下的增长速率高于黑麦草,而铜钱草则在3种营养水平下的增长速率均为最小。由图1(c)可知,群落3中美人蕉、鸢尾的增长速率在3种营养水平下均明显高于梭鱼草和铜钱草,美人蕉和鸢尾也均在低、高营养水平间存在显著性差异,但在低、中及中、高营养水平间的差异性不显著; 梭鱼草的增长速率虽随着营养水平的提高而增大,但在3种营养水平间的差异性不显著; 铜钱草则在中、高营养水平间的差异性不显著。由图1(d)可知,群落4中植物增长速率总体上与群落3相似,各植物均随营养水平的提高而加快增长,其中,美人蕉、鸢尾在3种营养水平下有着相对更好的表现,而梭鱼草次之,狐尾藻则在3种营养水平下的增长速率均表现不佳。
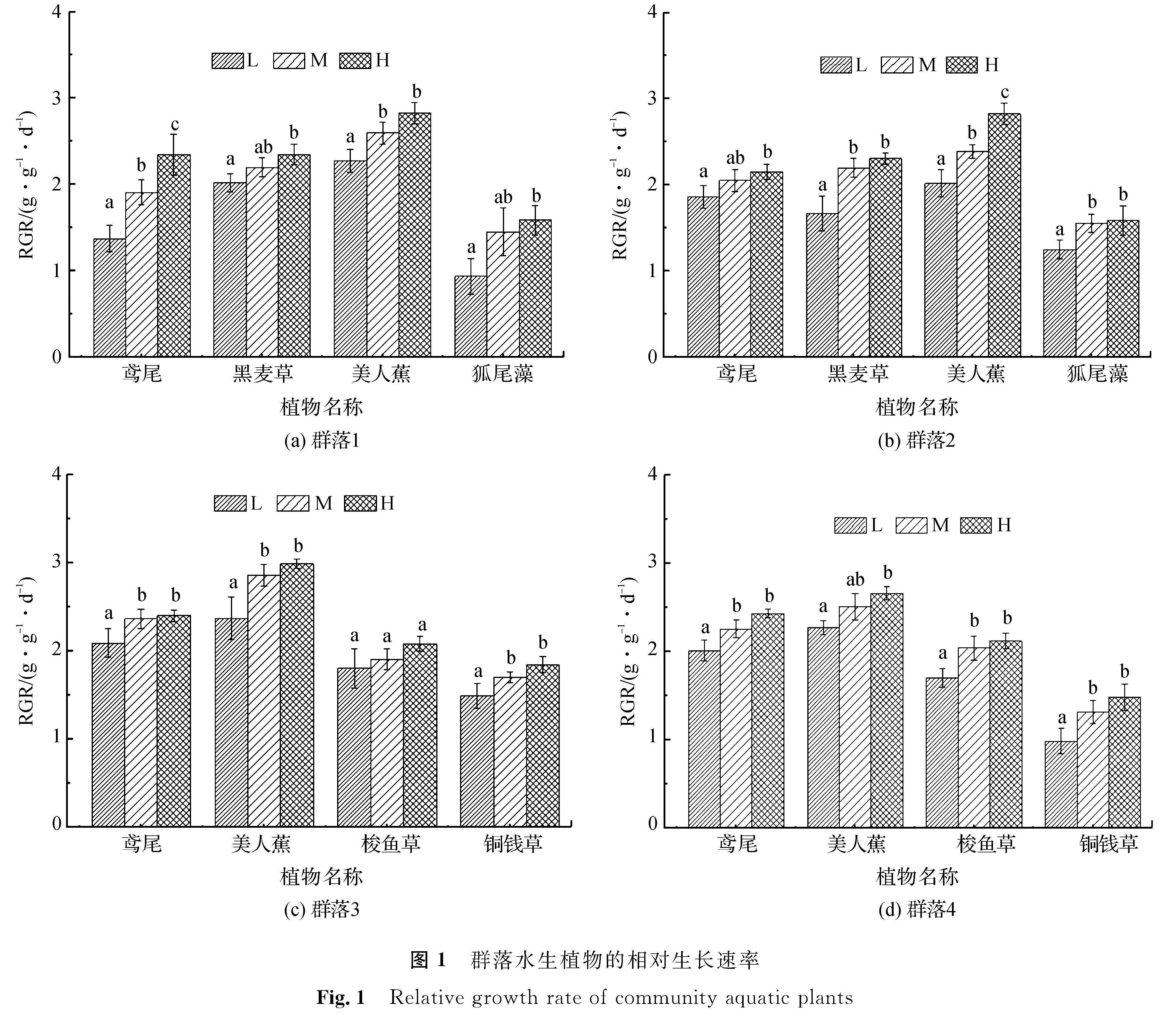
图1 群落水生植物的相对生长速率
Fig.1 Relative growth rate of community aquatic plants
2.2 群落中各水生植物氮、磷含量的变化分析
各群落中的水生植物在试验前后的氮含量变化见图2。由图2(a)可知,群落1中美人蕉氮含量变化值随营养水平的提高逐渐增大,但在低、中营养水平下的氮含量差异未达显著水平,而在高营养水平下的氮含量变化值为2.2 mg/g,与低、中营养水平下存在显著性差异,这可能意味着水体氮含量越高,美人蕉体内富集相对含量也越大。鸢尾、狐尾藻2种植物在低营养水平下氮含量变化为负,而在中、高营养水平下均略增加; 黑麦草则在3种营养水平下略增加,但变化幅度很小。对比图2(a)与(b)可知,群落2中植物的氮含量变化情况与群落1基本上一致,鸢尾和铜钱草两种植物在低营养水平下氮含量净变化为负,在中、高营养水平下则均为正; 黑麦草和美人蕉在3种营养水平下氮含量均有增加,但美人蕉明显比黑麦草在3种营养水平下的氮含量增加值更大,在高营养水平下氮含量变化值为1.9 mg/g。由图2(c)可知,群落3中鸢尾、美人蕉、铜钱草、铜钱草4种植物随营养水平的提高,氮含量均增大,且在低、高营养水平间均存在显著性差异; 梭鱼草氮含量变化在低、中及中、高营养水平间存在显著性差异,其余3种植物在中、高营养水平间则不存在显著性差异。由图2(d)可知,群落4中鸢尾、美人蕉在3种营养水平下的氮含量变化均明显高于梭鱼草和狐尾藻,而后两种植物在低营养水平下的氮含量变化为负。4种植物的相同点在于随营养水平的提高,氮含量变化值均有所增大。
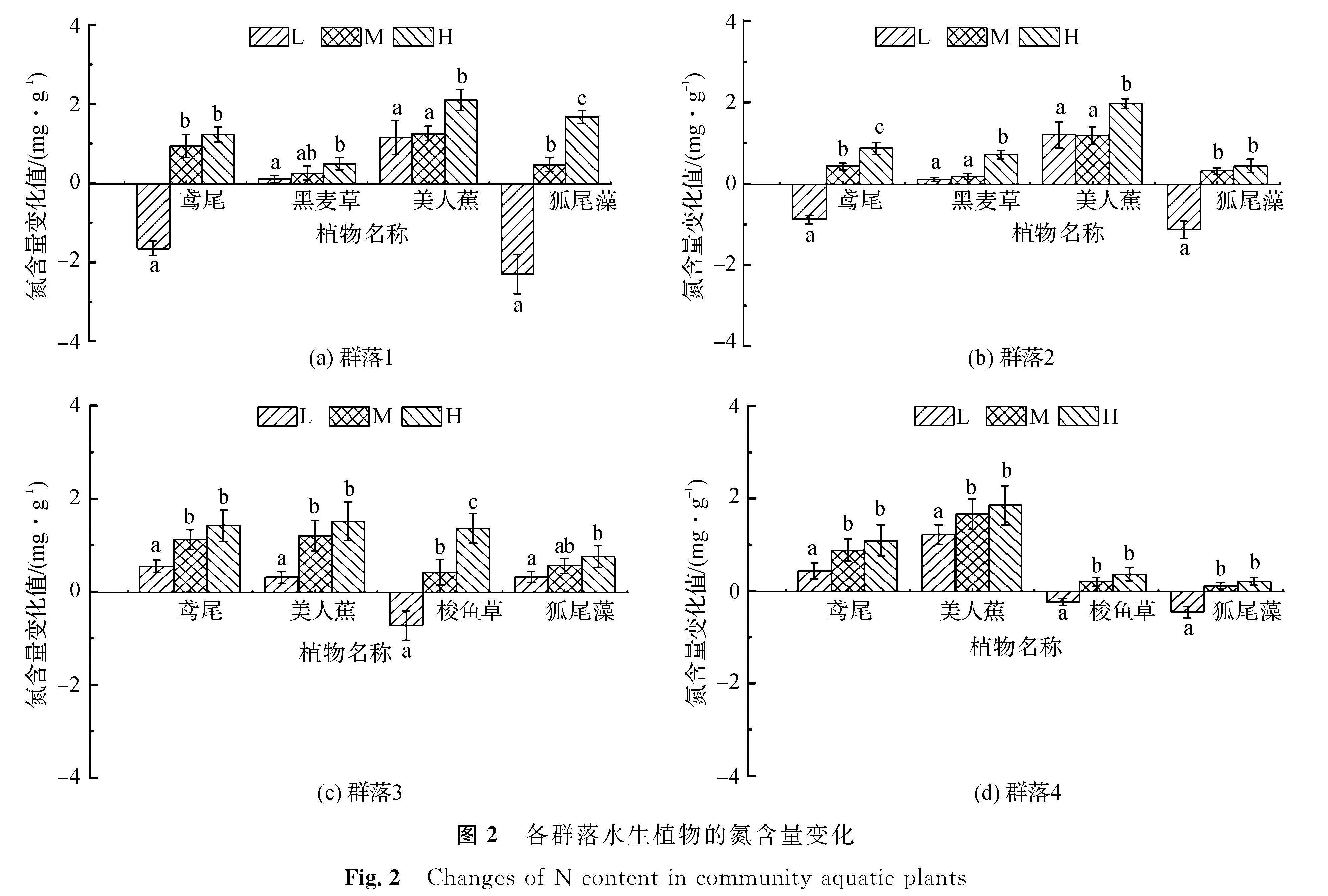
图2 各群落水生植物的氮含量变化
Fig.2 Changes of N content in community aquatic plants
各群落中的水生植物在试验前后的磷含量变化见图3。由图3(a)可知,群落1中4种植物的磷含量

图3 各群落水生植物的磷含量变化
Fig.3 Changes of P content in community aquatic plants
变化值均为正,且均随营养水平的提高而相对增大; 各植物在低、高营养水平间均存在显著性差异,但低、中营养水平间则均无显著性差异; 鸢尾、黑麦草、美人蕉在中、高营养水平间亦无显著性差异。由图3(b)可知,群落2中鸢尾、美人蕉、铜钱草3种植物在低营养水平下的磷变化值为负,中、高营养水平下则均为正; 鸢尾、美人蕉的磷含量变化幅度相对较大,而黑麦草、铜钱草则相对较小。由图3(c)可知,群落3中美人蕉、梭鱼草、铜钱草3种植物的磷变化值在各营养水平间均呈显著性差异,鸢尾在低、中营养水平间无显著性差异,但在低、高及中、高营养水平间均有显著性差异。由图3(d)可知,群落4中狐尾藻在低营养水平下磷含量变化值为负,在中、高营养水平下磷含量变化值为正。鸢尾与美人蕉在3种营养水平下的磷含量变化值大致相当,均略高于梭鱼草与狐尾藻。
2.3 群落中各水生植物氮、磷的积累群落植物体内氮、磷积累反映了植物对水中营养元素的富集能力,是水体生态化治理时选取植物的最重要指标。由于植物在不同群落中可能面临与其他植物之间的相互协同亦或是相互竞争作用,故需对比分析不同群落中各种水生植物在不同营养水平下的氮、磷积累状况,结果见图4~7。
从图4(a)可以看出,低营养水平下,群落1中各植物氮积累率由低至高依次为狐尾藻、鸢尾、黑麦草、美人蕉; 中营养水平下,各植物氮积累率由低至高依次为黑麦草、鸢尾、狐尾藻、美人蕉; 高营养水平下,各植物氮积累率由低至高依次为狐尾藻、黑麦草、鸢尾、美人蕉。鸢尾、美人蕉的氮积累率在3种营养水平间均呈显著性差异,在最高营养水平下氮积累率为3.6%。黑麦草、狐尾藻在低、高营养水平间氮积累率均呈显著性差异,但两植物在中营养水平下的氮积累率分别与低、高营养水平间无显著性差异。从总体上看,群落1中植物的氮积累率随营养水平的提高均有所增加。从图4(b)中可以看出,群落1中各植物的磷积累率随营养水平的提高均呈现相对增加的趋势,且各植物在3种营养水平下对应的磷积累率相对顺序保持不变,由低至高依次为狐尾藻、鸢尾、黑麦草、美人蕉。美人蕉在最高营养水平下的磷积累率为3.2%。美人蕉、鸢尾在3种营养水平间的磷积累率均呈显著性差异; 黑麦草则在低、中营养水平间磷积累率无显著性差异,狐尾藻在中、高营养水平间无显著性差异。
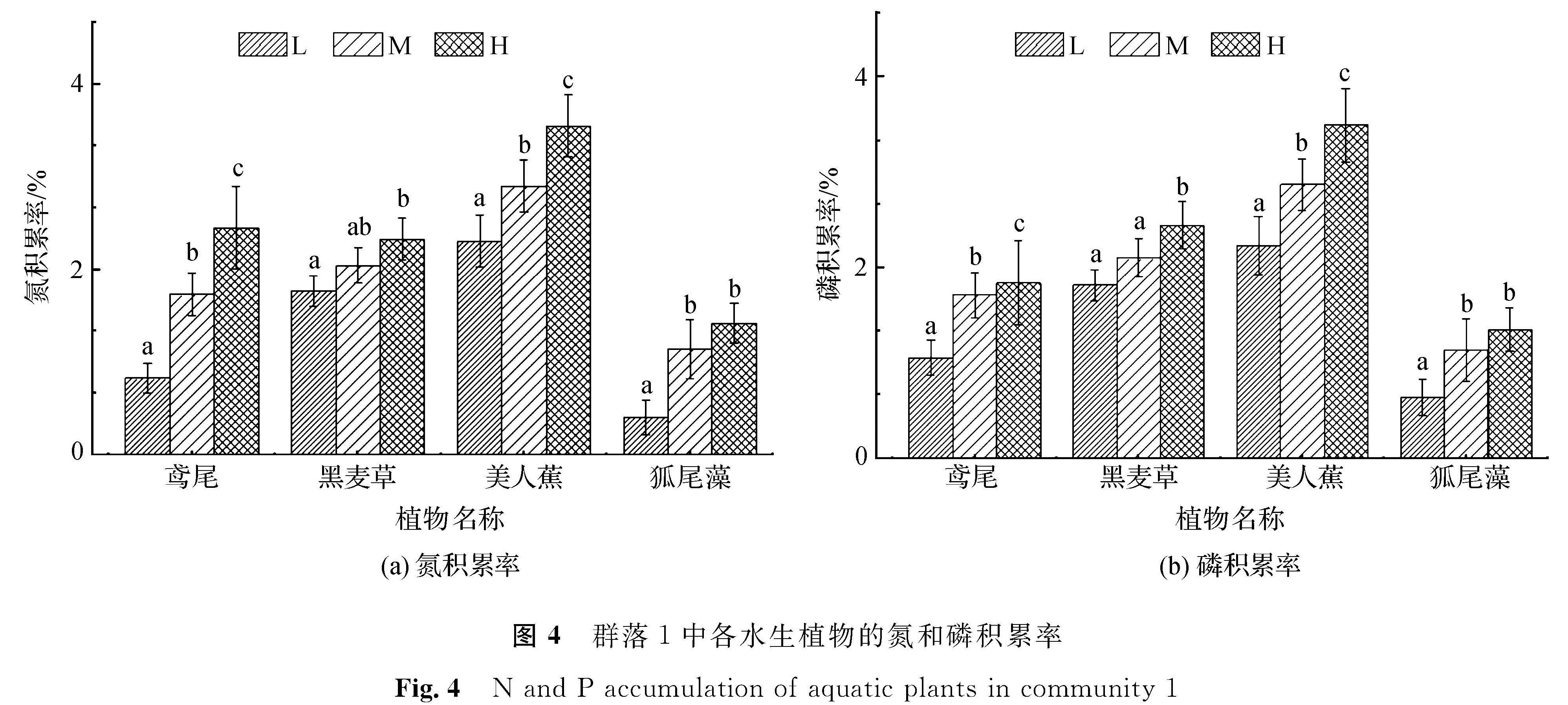
图4 群落1中各水生植物的氮和磷积累率
Fig.4 N and P accumulation of aquatic plants in community 1
从图5中可以看出,群落2中美人蕉在3种营养水平下的氮积累率均为最高,且层级间的氮积累率均呈显著性差异; 鸢尾在低营养水平下的磷积累率略高于黑麦草,但在中、高营养水平下的磷积累率则略低于黑麦草,而铜钱草则在3种营养水平下的磷积累率均为最低。群落2中美人蕉在3种营养水平下均保持相对最高的磷积累率; 鸢尾在低营养水平下的磷积累率高于黑麦草,但在中、高营养水平时略低于黑麦草; 铜钱草磷积累率则在3种层级下均为最低。从整体上看,群落2中植物的磷积累与氮积累情形大致相似。
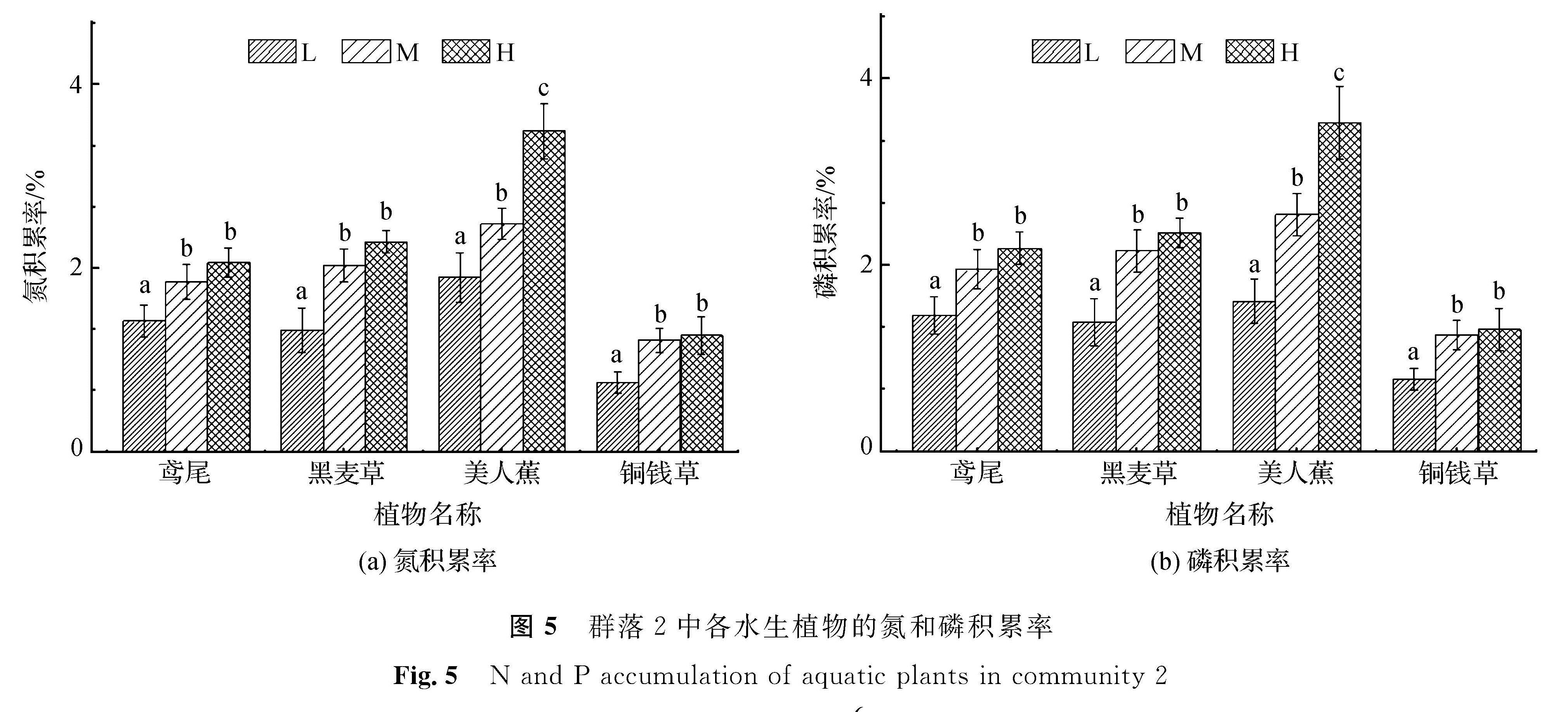
图5 群落2中各水生植物的氮和磷积累率
Fig.5 N and P accumulation of aquatic plants in community 2
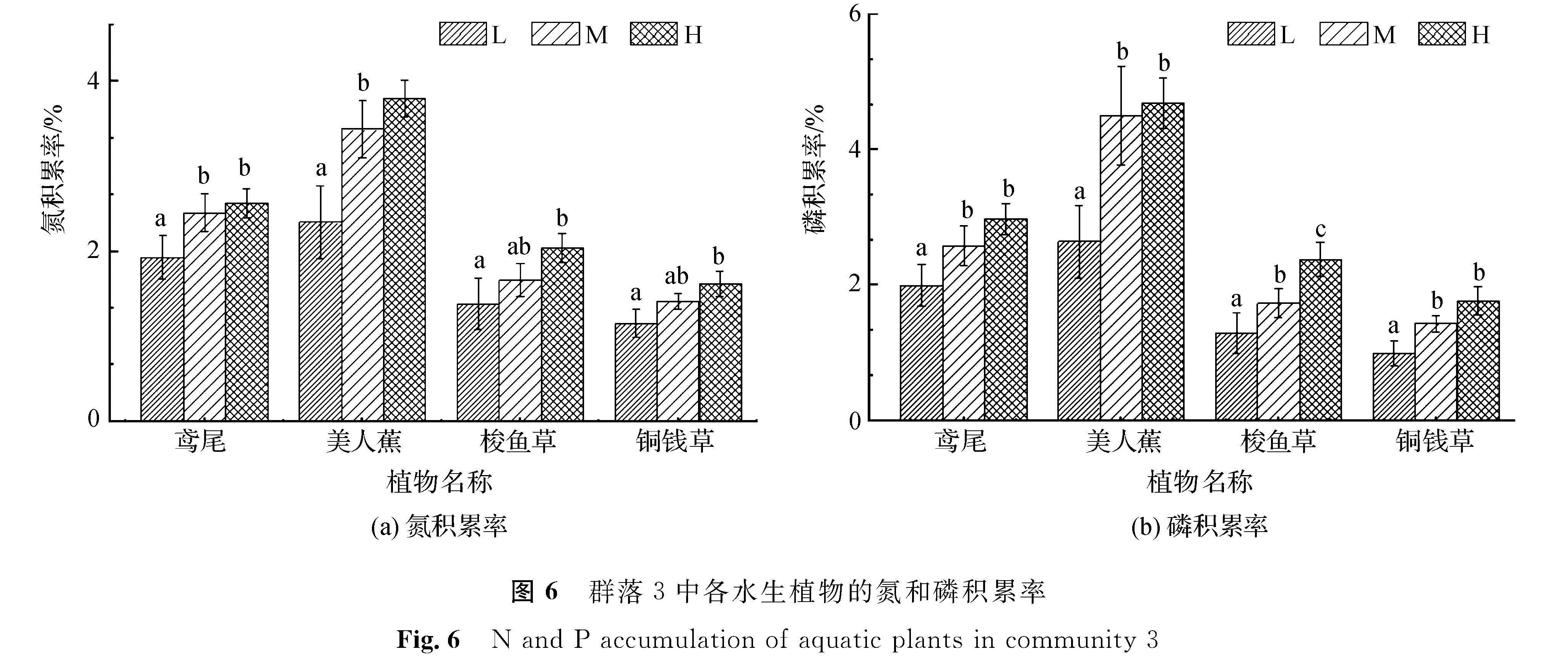
图6 群落3中各水生植物的氮和磷积累率
Fig.6 N and P accumulation of aquatic plants in community 3
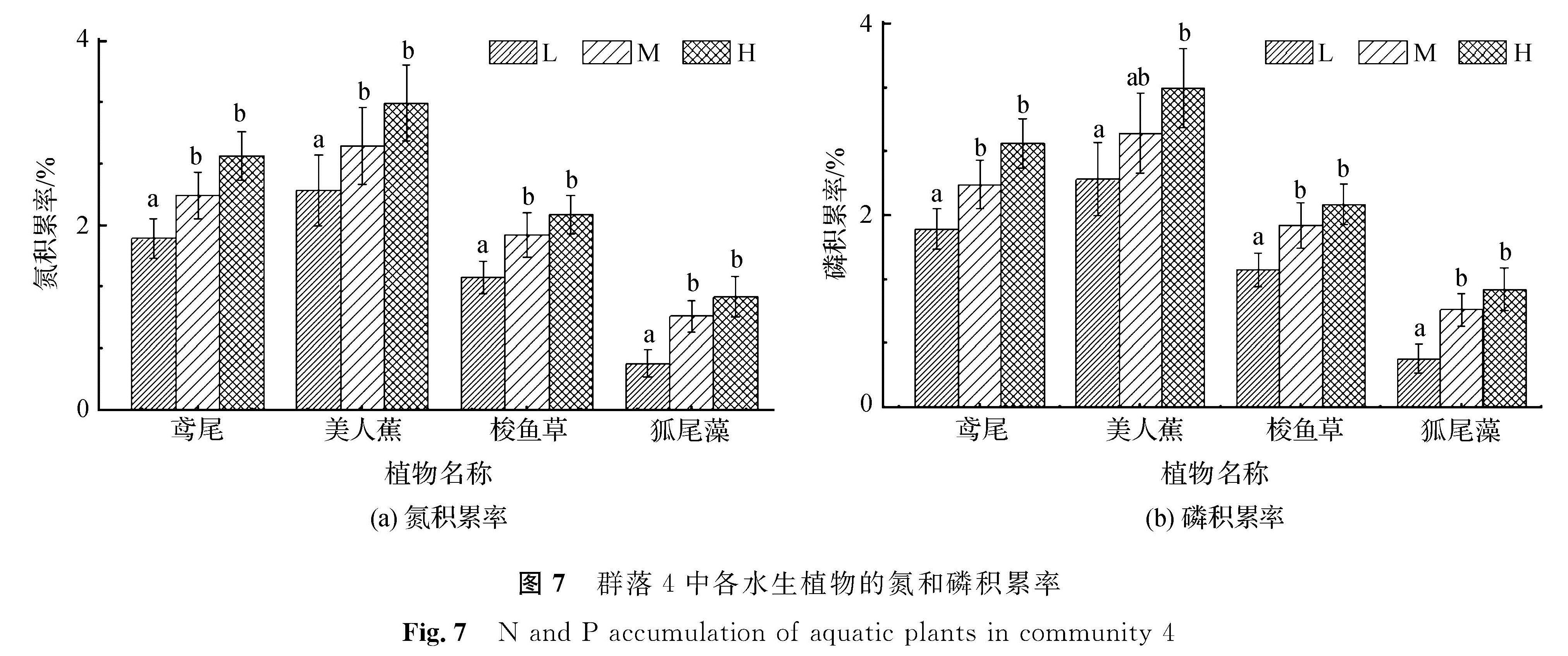
图7 群落4中各水生植物的氮和磷积累率
Fig.7 N and P accumulation of aquatic plants in community 4
从图6中可以看出,群落3中各植物的氮积累数值在3种营养水平下的相对关系保持一致,即在3种营养水平下氮积累率由低至高依次为铜钱草、梭鱼草、鸢尾、美人蕉。其中美人蕉在最高营养水平下的氮积累率为3.6%。各植物的氮积累率均在低、高营养水平间呈显著性差异,而在中、高营养水平间未出现显著性差异。鸢尾、美人蕉两种植物的氮积累率在低、中营养水平间存在显著性差异,而梭鱼草、铜钱草两种植物的氮积累率则在低、中营养水平间无显著性差异。群落3中,美人蕉在各营养水平的磷积累率均最高,最低者是铜钱草。鸢尾、美人蕉、铜钱草3种植物在中、高营养水平间无显著性差异,而在低、中及低、高营养水平间有显著性差异; 梭鱼草则在3种营养水平均存在显著性差异。
由图7可知,群落4中,狐尾藻氮积累率在3种营养水平下均为最低,而美人蕉氮积累率则在3种营养水平下均为最高。其中美人蕉在最高营养水平下的氮积累率为3.3%。4种植物的氮积累率均随营养水平的提高而增加,中、高营养水平间显著性差异不明显,而低、高营养层间的氮积累率存在显著性差异。群落4中,各植物在3种营养水平下的磷积累率高低依次为美人蕉、鸢尾、梭鱼草、狐尾藻。各植物的磷积累率随营养水平的提高而趋于增高,低、高营养水平间的磷积累率差异最为显著,而中、高营养水平间的差异不显著。
2.4 各群落整体氮、磷的积累各群落在不同营养水平下的氮、磷积累结果如图8所示。
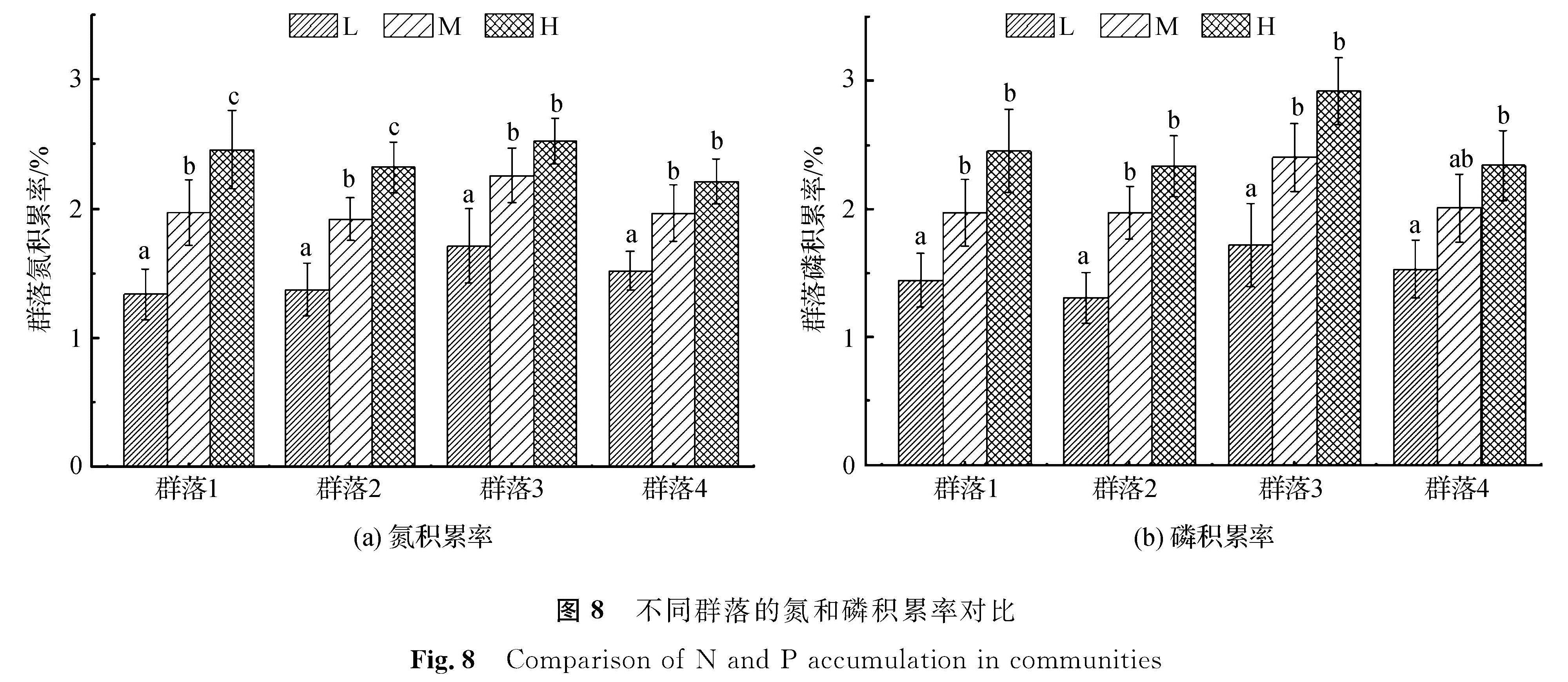
图8 不同群落的氮和磷积累率对比
Fig.8 Comparison of N and P accumulation in communities
由图8(a)可知,在低营养水平下,群落3的氮积累率最高,为(1.71±0.28)%,群落4次之,群落3略低于群落4,群落1的氮积累率最低,为(1.34±0.19)%; 在中营养水平下,群落3的氮积累率最高,为(2.26±0.21)%,群落1、群落2、群落4的氮积累率则大致相当; 在高营养水平下,群落3的氮积累率最高,为(2.52±0.17)%,群落1次之,群落2略低于群落1,群落4最低,为(2.21±0.17)%。群落1和群落2的氮积累率在两两营养水平间均表现出显著性差异,群落3和群落4的氮积累率在低、中及低、高营养水平间表现出显著性差异,在中、高营养水平间差异不显著。由图8(b)可知,低营养水平下,群落3的磷积累率高于其余3个群落,其值为(1.72±0.32)%。群落4的磷积累率则略高于群落1,群落2的磷积累率最低,为(1.31±0.20)%; 中营养水平下,群落3的磷积累率最高,为(2.40±0.26)%,群落1和群落2、群落4的磷积累率则大致相当; 高营养水平下,群落3的磷积累率依然最高,为(2.92±0.31)%,群落1的磷积累率次之,为(2.45±0.32)%,群落2和群落4的磷积累率则大致相当。各群落磷积累率在营养水平间的差异性与氮积累率大致相近,表现为低、中及低、高营养水平间差异性显著,中、高营养水平间的差异未出现显著性差异。
3 结 语本试验结果表明,植物在群落中的生长状况与其在单独种植时的情生长状况存在差异,群落中植物的氮、磷含量也与其单独种植时有所差异,相应的群落中植物的氮、磷积累与其单独种植时也有所不同。由于植物氮、磷含量的变化值较其含量值而言小得多,因此植物氮、磷积累的主导因素是植物的生物量,这意味着植物的生长速率对其氮、磷积累起主导作用。植物单独种植及在群落中种植时的差异,也反映出群落植物间存在相互竞争与协同的关系。这种关系不仅与植物类型有关,同时也与环境条件有关,包括营养元素水平、微量元素水平、水温、光照及水体交换速度等因素。就试验结果而言,群落3(植物组合为鸢尾+美人蕉+梭鱼草+铜钱草)对氮、磷均有相对更高的积累率,故其用于富营养化水体的生态化治理更为适宜。
- [1] 陈巧玲,林晓葱,宫本涛,等.12种水生植物对氨氮和总磷的净化效果研究[J].福建农业科技,2019(1):45.
- [2] 易旺,宋建军,邓洁,等.水生植物对富营养化水体的净化作用及其影响因素综述[J].南方农业,2019,13(32):174.
- [3] 刘敏,吴铁明,刘菡,等.3种水生植物的不同组合对富营养水体的净化效果研究[J].中国农业科技导报,2019,21(7):157.
- [4] 周晓燕,贾涛,杨涓.几种水生植物去污能力比较研究[J].安徽农学通报,2017,23(21):36.
- [5] 张芳,易能,张振华,等.不同类型水生植物对富营养化水体氮转化及环境因素的影响[J].江苏农业学报,2015,31(5):1047.
- [6] 张力,张振华,高岩,等.不同水生植物对富营养化水体释放气体的影响[J].生态与农村环境学报,2014,30(6):740.
- [7] 洪瑜,王英,王芳,等.不同水生植物组合对稻田退水的氮磷净化效果[J].环境科学与技术,2014,30(6):736.
- [8] 倪洁丽,王微洁,谢国建,等.水生植物在水生态修复中的应用进展[J].环保科技,20016,22(3):43.
- [9] 郑足红,胡超,王华伟,等.3种水生植物对富营养化水质的净化性能比较[J].环境污染与防治,2018,40(10):1148.
- [10] 聂司宇,孟昊,李婷婷,等.水生植物对富营养化水体中氮磷去除的研究进展[J].环境保护与循环经济,2020,40(4):49.
- [11] 陈远帆.生物种间关系的复杂性[J].生物学杂志,1993(2):46.
- [12] 谭凯婷,柳君侠,王志红,等.沉水植物修复富营养化景观水体的研究进展[J].水处理技术,2019,45(6):17.
- [13] 刘建伟,周晓,吕臣,等.三种挺水植物对富营养化景观水体的净化效果[J].湿地科学,2015,13(1):10.
- [14] 李欢,吴蔚,罗芳丽,等.4种挺水植物、4种沉水植物及其组合群落去除模拟富营养化水体中总氮和总磷的作用比较[J].湿地科学,2016,14(2):168.
- [15] 袁杰,董立新,杨洁,等.六种挺水植物对富营养化河水氮磷净化效果研究[J].环境科学与管理,2017,42(4):78.
- [16] 谭凯婷,柳君侠,王志红,等.沉水植物修复富营养化景观水体的研究进展[J].水处理技术,2019,45(6):17.
- [17] 窦艳艳,陈垚,李春龙.不同水生植物组合对富营养化水体净化的模拟研究[J].环境科学与技术,2015,38(8):272.
- [18] 周玥,韩玉国,张梦,等.4种不同生活型湿地植物对富营养化水体的净化效果[J].应用生态学报,2016,27(10):3354.
- [19] 张倩妮,陈永华,杨皓然,等.29种水生植物对农村生活污水净化能力研究[J].农业资源与环境学报,2019,36(3):394.
- [20] 国家环境保护总局.水质 总氮的测定 碱性过硫酸钾消解紫外分光光度法:HJ 636—2012[S].北京:中国环境科学出版社,2012.